Разложение соломы озимой пшеницы с помощью микромицетов-целлюлозолитиков
Безлер Н. В., кандидат сельскохозяйственных наук, зав. лабораторией эколого-микробиологических исследований почвы, Колесникова М. В., аспирант, Всероссийский НИИ свеклы и сахара им. А. Л. Мазлумова
Значительный ущерб окружающей среде наносит сжигание соломы, оставшейся после уборки зерновых культур, доля которой составляет 50 — 70% от их урожая. Существенные затраты идут на уборку этого малоценного в кормовом отношении продукта (50 — 65%). Излишки соломы часто не убирают, а сжигают на месте. Дым от палов — мощный загрязнитель атмосферного воздуха. Каждые 1000 га сожженной стерни и соломы выделяют в атмосферу 500 кг окислов азота, 37 кг углеводородов, 20 т углекислого и угарного газов, а также 3 т золы. Закись азота и диоксид углерода являются парниковыми газами, и сокращение поступления их в атмосферу — важная задача на современном этапе развития производства.
Вместе с растительными остатками огнем уничтожаются и целые биоценозы. Потеря гумуса в верхнем слое почвы при сжигании пожнивных остатков составляет 1,3 т/га, что приводит к снижению плодородия почвы.
В то же время солома является важным источником пополнения запасов углерода в почве, служит материалом для образования гумуса. При ее запашке в почву возвращается (в пересчете на 1 га) 12—15 кг азота, 7 — 8 кг фосфора, 24 — 30 кг калия. Разложение соломы, как и любого, не содержащего азот, вещества, требует дополнительного поступления этого элемента питания в почву. Расчетный расход азота составляет 0,5—1,3% от массы соломы, то есть 40 — 80 кг на 1 га. В связи с этим, существует проблема утилизации излишка соломы таким образам, чтобы продукты ее разложения участвовали в синтезе гумуса.
Главными потребителями целлюлозы, из которой на 70% состоит солома зерновых культур, в почве являются микромицеты. Численность их в черноземах невелика, к тому же невысокое содержание влаги в почве не способствует быстрому разложению соломы. Штаммы микромицетов, выделенные из черноземов и обладающие высокой целлюлозолитической активностью, способны ускорить этот процесс.
В лаборатории эколого-микробиологических исследований почвы ВНИИСС в коллекции эффективных микроорганизмов культивируются микромицеты-целлюлозолитики. Из них мы выбрали штамм, способный разлагать солому и синтезировать меланины, которые входят в состав гумуса. В результате исследований был отобран лучший штамм под индексом 0, (рис. 1).
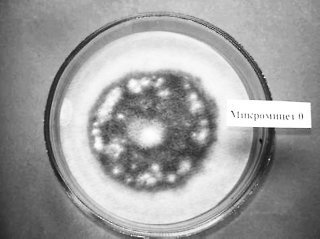
Рис. 1. Микромицет-целлюлозолитик, развитие на фильтровальной бумаге
В лабораторных условиях мы установили режимы разложения соломы озимой пшеницы с помощью выделенного аборигенного штамма микромицета чернозема выщелоченного, обладающего высокой целлюлозолитической активностью.
Опыты по установлению целлюлозолитической активности микромицета были заложены в моделируемых условиях, в четырехкратной повторности. Режим культивирования был близким по температурно-влажностным показателям, складывающимся в конце августа-сентября в почвах ЦЧР.
Результаты исследования показали, что без участия микромицета не отмечено существенного разложения соломы. В присутствии минерального азота этот процесс несколько активизируется. Введение в субстрат микромицета с питательной добавкой (ПД) увеличило степень разложения соломы до 15%. Состав питательной добавки является ноу-хау авторов. Интенсивное разложение соломы отмечено при совместном применении микромицета (4300 шт. спор в 1 мл), питательной добавки и азота, где процент ее разложения колеблется от 55% до 75%, (Рис. 2).

Рис. 2. Разложение соломы под влиянием микромицета-целлюлозолитика (вариант 1—солома, обработанная водой; вариант 16 — солома, обработанная микромицетом-целлюлозолитиком с азотом и ПД)
Таким образом, на процесс разложения соломы озимой пшеницы влияет доза микромицета-целлюлозолитика, а также совместное его применение с питательной добавкой и азотом, которые стимулируют его рост, и тем самым способствуют ускорению деструкции соломы.
Одновременно мы определяли токсичность полученного субстрата, в качестве тест-культуры применяли семена редиса. Исследования показали, что в контроле, в качестве которого использовали фильтровальную бумагу, смоченную дистиллированной водой, длина корешка составила 22,5 см. Длина корешка тест-культуры, проросшей на чистой соломе, равнялась 10,9 см, а в варианте, где была отмечена максимальная степень разложения соломы — 13,6 см. Это свидетельствует о том, что под влиянием микромицета отмечается тенденция к снижению токсичности соломы.
На основании результатов лабораторных исследований, был заложен полевой опыт на новом опытном поле ВНИИСС на фоне без удобрений. Площадь делянки 27 м2. Расположение делянок — систематическое, в четырехкратной повторности. Почва участка — чернозем выщелоченный среднесуглинистый среднегумусный со средним содержанием элементов питания, pH = 6,2. Для посева использовали семена гибрида сахарной свеклы РМС 70.
Технология возделывания сахарной свеклы была общепринятой для ЦЧР.
По мере роста и развития сахарной свеклы, проводили наблюдения за изменениями, происходящими в микробном сообществе чернозема выщелоченного. Численность важнейших таксономических (микромицеты, бактерии и актиномицеты) и экологотрофических групп микроорганизмов учитывали методом высева почвенной суспензии различной степени разведения на элективные питательные среды. («Методы почвенной микробиологии и биохимии». Под ред. Д. Г. Звягинцева — М., Изд-во МГУ, 1991; Е. З. Теппер, В. К. Шильникова, Г. И. Переверзева «Практикум по микробиологии», 2004.)
Внесение в почву соломы увеличило от начала до конца периода вегетации численность микроорганизмов, использующих минеральные формы азота. На контроле численность микроорганизмов этой группы снизилась к концу вегетационного периода на 1,3 млн КОЕ в 1 г абсолютно сухой почвы. Добавление к соломе азота повысило их численность на 2,3 млн КОЕ в начале вегетации и на 16,5 млн КОЕ — в конце вегетации по сравнению с контролем, что вполне закономерно при поступлении в почву дополнительного количества минерального азота. Питательная добавка активизировала минерализационные процессы в почве к концу вегетационного периода и увеличила к этому моменту численность микроорганизмов, использующих минеральные формы азота, в 3 раза по сравнению с контролем. Применение микромицета с питательной добавкой и азотом повысило численность микроорганизмов этой группы, по сравнению с контролем, в мае — на 1,9 млн КОЕ, в сентябре — на 15,6 млн КОЕ в 1г абсолютно сухой почвы.
Численность аммонификаторов на контроле к середине вегетационного периода увеличилась на 2,5 млн КОЕ, но к концу вегетации осталась той же — 3,9 млн КОЕ в 1 г сухой почвы. Внесение одной соломы снизило в мае их численность на 1,0 млн. КОЕ по сравнению с контролем, а к сентябрю наметилась тенденция к повышению численности этой группы микроорганизмов. Добавление к субстрату минерального азота увеличило численность аммонификаторов в начале и в конце периода вегетации, по сравнению с контролем, в мае — на 17-18%, а в сентябре — на 20%. Под действием питательной добавки численность аммонификаторов в начале вегетации возросла на 0,3 млн КОЕ, в конце— на 1,4 млн КОЕ. Взаимодействие микромицета-целлюлозолитика с почвенной микрофлорой увеличило численность аммонификаторов на первых этапах вегетации на 26% по сравнению с контролем. Это свидетельствует о более высокой интенсивности минерализационных процессов под его влиянием (таблица 1).
|
Таблица 1 Влияние микромицета и ПД на минерализационные процессы в почве под сахарной свеклой (млн КОЕ в 1 г сухой почвы)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
С увеличением численности аммонификаторов начинается активная минерализация свежего органического вещества (идет накопление доступных форм элементов питания). Основу для этих процессов дает субстрат, а именно, солома озимой пшеницы.
Другой важный процесс, протекающий в почве, — синтез и распад гумуса. Зимогенная микрофлора играет роль поставщика материалов, из которых синтезируются молекулы гумуса и ферментов, принимающих участие в этом процессе. В противоположном процессе (деструкции гумуса) принимает участие автохтонная микрофлора. На протяжении всего периода вегетации увеличила численность зимогенная микрофлора. На фоне минерального азота численность зимогенной микрофлоры была значительно выше, чем в других вариантах, как в начале, так и к концу вегетации. Ее численность была выше в мае на 2,6 млн КОЕ, а в сентябре — на 17,6 млн КОЕ по сравнению с контролем. Внесение вместе с соломой в почву микромицета повысило численность этой группы почвенной микрофлоры в середине вегетационного периода на 1,2 млн КОЕ, а в конце вегетации — на 15,1 млн КОЕ по сравнению с контролем (таблица 2).
|
Таблица 2 Динамика гумификационных процессов в почве при внесении соломы (млн КОЕ в 1 г абсолютно сухой почвы)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Численность автохтонной микрофлоры повышается с начала вегетационного периода и до его окончания с 9,2 до 11,7 млн КОЕ в 1 г абсолютно сухой почвы. При внесении соломы в почву численность автохтонной микрофлоры снизилась в мае до 5,37 млн КОЕ, а к середине вегетации повысилась до 7,27 млн КОЕ и до сентября удерживалась практически на одном уровне.
Введение в субстрат микромицета увеличивало численность микроорганизмов этой группировки на протяжении всего периода вегетации.
Под влиянием совместного внесения в почву микромицета-целлюлозолитика с соломой усиливаются гумификационные процессы в почве. Так, в начале вегетационного периода на контроле К гумификации составил 0,38, при внесении соломы — 0,23, а при использовании микромицета — 0,75. Таким образом, интенсивность синтеза гумуса повысилась в 2 раза. В середине вегетационного периода гумификация увеличилась на 4%. По сравнению с чистой соломой, скорость гумификации в этом варианте на первых этапах вегетационного периода увеличилась в 3 раза. К концу вегетационного периода интенсивность процессов гумификации возрастает, однако направленность процесса по вариантам изменяется. Пик этого процесса перемещается к варианту с чистой соломой. Это связано с ее ускоренным разложением под воздействием микромицета-целлюлозолитика и усилением гумификационных процессов в начале вегетации.
Микробиологическая деятельность в почве в сильной степени ограничивается из-за отсутствия достаточного количества свежего органического вещества. При обогащении почвы соломой резко увеличивается количество микроорганизмов, разлагающих сложные полимерные соединения.
Наличие соломы в почве существенно не повлияло на численность микроскопических грибов в начале вегетационного периода. Исключением явилось внесение с соломой микромицета-целлюлозолитика. В этом случае численность микроскопических грибов увеличилась, по сравнению с контролем, с 13,9 до 16,4 тыс. КОЕ в 1 г абсолютно сухой почвы (таблица 3).
|
Таблица 3 Влияние микромицета и ПД на численность микроорганизмов, разлагающих сложные полимерные соединения в почве (в 1 г абсолютно сухой почвы)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Особенно сильное различие наблюдалось в середине вегетации, когда численность микромицетов повысилась с 12,4 до 29,7 тыс. КОЕ в 1 г абсолютно сухой почвы соответственно. К концу вегетации наибольшая численность микромицетов наблюдалась при внесении чистой соломы (60,9 тыс. КОЕ в 1 г абсолютно сухой почвы) совместно с микромицетом-целлюлозолитиком, ПД и азотом — 50,3 тыс. КОЕ в 1 г абсолютно сухой почвы, что выше контроля на 17,3 и 3,5 тыс. КОЕ в 1 г абсолютно сухой почвы соответственно. Таким образом, микромицет-целлюлозолитик способствует повышению общей численности микромицетов в почве. Однако она не превышает оптимальных границ.
В тех же условиях внесение соломы озимой пшеницы стимулировало развитие актиномицетов к концу периода вегетации. Добавление азота к субстрату увеличило их численность в сентябре в 3 раза по сравнению с контролем. Действие питательной добавки повысило численность актиномицетов на 2,4 млн КОЕ в 1 г абсолютно сухой почвы по сравнению с контролем. Совместное использование микромицета, питательной добавки и азота способствовало увеличению численности актиномицетов в начале периода вегетации на 0,5 и в конце вегетации — на 0,7 млн КОЕ в 1 г абсолютно сухой почвы по сравнению с контролем.
Наличие субстрата (соломы) в почве определяет количество целлюлозоразрушающих микроорганизмов. Внесение соломы без каких-либо добавок привело к увеличению численности целлюлозолитиков в середине периода вегетации на 0,4 млн шт. в 1 г абсолютно сухой почвы. Питательная добавка увеличила их численность в июле на 1,6 млн КОЕ, а в сентябре — на 1,0 млн КОЕ в 1 г абсолютно сухой почвы по сравнению с контролем. Добавление к субстрату микромицета с питательной добавкой стимулировало развитие целлюлозолитиков. В начале периода вегетации их численность была выше контроля на 0,3 млн штук. К середине вегетации численность целлюлозоразрушающих микроорганизмов несколько снизилась, но превосходила контроль в 2 раза. К концу вегетации их численность увеличилась и была выше по сравнению с другими вариантами.
Совместное использование микромицета, питательной добавки и азота увеличило численность целллюлозолитиков в конце вегетации на 85% по сравнению с контролем. Это свидетельствует о том, что в почве активизировались процессы разложения соломы.
Таким образом, внесение соломы и микромицета-целлюлозолитика в почву, а также его совместное взаимодействие с питательной добавкой и азотом способствуют повышению активности полезной микрофлоры почвы, что стимулирует процессы, формирующие эффективное и потенциальное плодородие почвы, рост и развитие сахарной свеклы.
Погодные условия в начале вегетационного периода отличались недостатком влаги. Однако осадки, выпавшие после появления первых всходов сахарной свеклы, создали благоприятные условия для роста растений и прорастания остальных семян (таблица 4).
Влияние микромицета и ПД на динамику всходов сахарной свеклы
|
Таблица 4
|
|||||||||||||||||||||||||||||||||||||||||||||||
Наблюдения за динамикой появления всходов показали, что микромицет-целлюлозолитик ускорил появление всходов. Их густота увеличилась на 2,0 — 3,2 шт. на 1 погонный метр. За весь период наблюдений лучшие результаты были получены при совместном использовании микромицета-целлюлозолитика с питательной добавкой и азотом. Густота всходов заметно увеличилась и в последний день исследования имела самый высокий показатель — 22,5 проростков на 1 п.м., а на контроле—19,3 проростков на 1 погонный метр.
В фазе первой пары листьев определяли влияние внесенного в почву микромицета на распространение корнееда и на массу 100 растений (таблица 5).
|
Таблица 5 Влияние микромицета-целлюлозолитика и ПД на массу растений и на распространение корнееда
|
||||||||||||||||||||||||||||||||||||||||
Исследования показали, что совместное применение микромицета с питательной добавкой и азотом повысило массу 100 растений на 1,6%. Распространение заболевания корнеедом составило 17,6% (на контроле — 21%).
В других вариантах масса 100 растений ниже, чем на контроле. Это свидетельствует о повышении токсичности почвы при внесении соломы без микромицета-целлюлозолитика, что согласуется с результатами, полученными в лабораторных опытах.
Количество растений, заболевших корнеедом, в опыте снизилось по сравнению с контролем (21,0%). В варианте солома и азот распространение заболевания составило 12,3%, солома +ПД + N (40 кг д.в./га)+ микромицет—17,3%.
Итоговым показателем, характеризующим ростовые процессы, являются урожайность и сахаристость корнеплодов сахарной свеклы (таблица 7).
Внесение соломы в различных сочетаниях с азотом, ПД и микромицетом-целлюлозолитиком, по сравнению с контролем, повысили урожайность и сахаристость корнеплодов сахарной свеклы. При внесении соломы наметилась тенденция увеличения продуктивности сахарной свеклы: прибавка урожайности составила 2,6 т/га, сбор сахара увеличился на 5,9 т/га.
|
Таблица 7 Влияние микромицета-целлюлозолитика и ПД на продуктивность сахарной свеклы
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Добавление азота к соломе достоверно повысило урожайность сахарной свеклы на 11,0 т/га и сахаристость — на 0,3%, а сбор сахара повысился на 1,9 тонн на гектар. Действие питательной добавки увеличило продуктивность корнеплодов на 3,8 т/га, сахаристость — на 0,3%, сбор сахара — на 0,7 т/га. Совместное применение микромицета-целлюлозолитика с питательной добавкой и азотом достоверно увеличило урожайность сахарной свеклы на 12,5 т/га, сахаристость — на 0,4% и сбор сахара — на 2,2 тонны на гектар.
Преимущество совместного использования микромицета-целлюлозолитика с азотом и питательной добавкой, по сравнению с внесением азота и соломы, а также чистой соломы, заключается в том, что, благодаря применению микромицета, усиливаются гумификационные процессы в почве в первой половине вегетации, снижается ее токсичность, вызванная внесением соломы, увеличивается масса 100 растений, которые дополнительно получают стимуляцию ростовых процессов.
Таким образом, предварительные исследования показали, что целесообразно запахивать солому совместно с микромицетом-целлюлозолитиком, питательной добавкой и азотом, что способствует ускорению разложения соломы и сохранению плодородия почвы, сокращая загрязнение окружающей среды. При этом продуктивность сахарной свеклы повысилась на 37,4%.
Статья из брошюры «Биологические препараты. Сельское хозяйство. Экология: Практика применения»
Скачать брошюру в PDF-формате
Купить ЭМ-препараты в каталоге АРГО:
- Байкал ЭМ-1
- ЭМ-патока
- Тамир, концентрат
- Ургаса
- ГуматЭМ Универсальный
- ГуматЭМ для корневой и внекорневой подкормки всех видов культур и замачивания семян
- ГуматЭМ для плодово-ягодных, цветочно-декоративных многолетних культур
- ГуматЭМ для защиты растений
- ЭМ-Культиватор
- Концентрат кормовой «ЭМ-курунга» ВетЭМ
- ЭМ-контейнер
- Биодезодорант бытовой «Эмикс «Чистый воздух»
- ЭМ-пластина
- ЭМ-доска, подставка вкуса
- ЭМ-керамика
- ЭМ-Курунга (БАДы)
Товары из статьи:

